ЛЕКЦІЯ № 9 3 страницаЗалежно від умов функціонування автономна нервова система може здійснювати на органи коригувальний і пусковий вплив. Коригувальний вплив полягає у тому, що коли орган, володіючи автоматією, функціонує, то імпульси, які надходять автономними нервами, тільки посилюють чи послаблюють його діяльність. Якщо ж робота органа не є постійною, а збуджується імпульсами, що надходять симпатичними чи парасимпатичними нервами, має місце пусковий вплив. Найчастіше пускові впливи доповнюють коригувальні. 8.2. Вплив фармакологічних засобів на функції автономної нервової системи
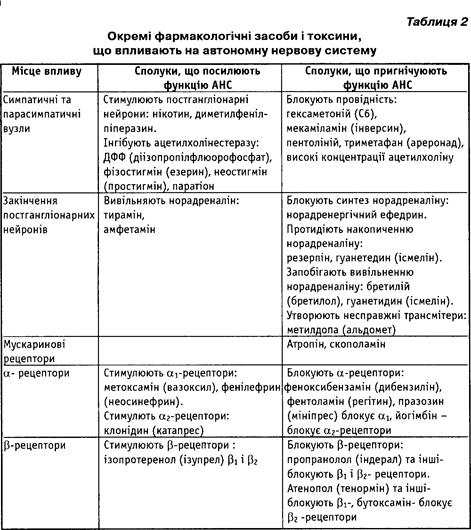
Синаптичні контакти рухових шляхів периферійного відділу автономної нервової системи є місцем дії фармакологічних препаратів на вісцеральні функції, оскільки збудження у цих контактах передається за допомогою хімічних нейротрансмітерів. Нейротрансмітери синтезуються, накопичуються у нервових закінченнях і вивільняються, діючи на нейрони, м'язові клітини або клітини залоз. Вплив на рецептори цих клітин ініціює їхні специфічні функції. Хімічні трансмітери усуваються шляхом поглинання або метаболізуються. Кожен з цих етапів може бути стимульованим або інгібованим із передбачуваним наслідком. У норадренергічних нервових закінченнях певні фармакологічні засоби зумовлюють утворення сполук, які заміщують норадреналін, і ці "несправжні нейротрансмітери" починають вивільнятися під впливом потенціалу дії, що надходить до нервового закінчення. Окремі фармакологічні засоби і токсини, які впливають на синаптичні контакти автономної нервової системи, можуть підсилювати або пригнічувати її функцію (табл. 2). Сполуки, що діють мускариноподібно, - це речовини, споріднені з ацетилхоліном, а також ті (засоби), що інгібують ацетилхолінестеразу. До таких засобів належать компоненти нервово-паралітичних газів (діізопропілфлюорофосфат - ДФФ) та інсектицид паратіон, які зумовлюють загибель організму шляхом масивного інгібування ацетил-холінестерази.
8.3. Структурні особливості автономної нервової системи
Підґрунтям функціональної діяльності автономної нервової системи є принцип рефлекторного регулювання, особливості якого визначаються структурними характеристиками системи. Основні відмінності автономної нервової системи від соматичної: 1) локалізація ядер у ЦНС; 2) мала величина нейронів; 3) опосередкований вихід волокон з мозку і відсутність чіткої сегментації їх поділу на периферії; 4) наявність автономних гангліїв на периферії; 5) еферентні волокна, які спрямовуються з мозку до внутрішніх органів, обов'язково перериваються у гангліях, де утворюють синапси на нейронах, що розташовані у цих гангліях; 6) безпосередній вихід на внутрішні органи. Автономна нервова система залежно від свого функціонального призначення поділяється на симпатичний (рис. 26) і парасимпатичний (рис. 27) відділи. Функціонально виділяють ще й третій відділ -метасимпатичний. Між симпатичним і парасимпатичним відділами є не
лише функціональні, але й структурні відмінності: а) за локалізацією центрів у мозку; б) за розташуванням гангліїв: ганглії парасимпатичного відділу розташовані в іннервованих ними органах (інтрамурально) або поруч із ними, тоді як ганглії симпатичного відділу локалізовані в симпатичному прикордонному стовбурі.
Центри автономної нервової системи
Скупчення нейронів, що утворюють автономні центри, локалізуються у мозковому стовбурі і спинному мозку. Центри парасимпатичного відділу локалізуються: 1) у середньому мозку (мезенцефальний відділ): автономні волокна йдуть у складі окорухового нерва; 2) у довгастому мозку (бульбарний відділ): еферентні волокна йдуть у складі лицевого,
відділ). Автономні волокна від них виходять через передні корінці спинного мозку разом із відростками мотонейронів. Центри автономної нервової системи перебувають у стані постійного тонусу, внаслідок чого іннервовані ними органи постійно отримують збуджувальні або гальмівні імпульси. Основне значення у забезпеченні цього тонусу належить аферентним нервовим сигналам, що надходять від рецепторів внутрішніх органів і, зокрема, від екстерорецепторів. Важливим є також вплив на центри автономної нервової системи різноманітних факторів крові і спинномозкової рідини. В органах з подвійною іннервацією (симпатичною і парасимпатичною) у стані фізіологічного спокою переважає вплив парасимпатичного нерва. Так, під дією імпульсів, що надходять до серця через блукаючий нерв, знижується частота і сила серцевих скорочень. Усі рівні системи підпорядковуються вищим підкірковим автономним центрам, які розташовані у відділі проміжного мозку - гіпоталамусі. Ці центри координують функції багатьох органів і систем організму, регулюючи тонус центрів мозкового стовбура і спинного мозку. Вони, у свою чергу, підпорядковуються корі великого мозку, що і забезпечує цілісне реагування організму із поєднанням його соматичних і автономних функцій в єдиний поведінковий акт. Двонейронна структура автономної рефлекторної дуги. На відміну від соматичної нервової системи, еферентний шлях якої до виконавчих органів однонейронний, периферійна частина всіх еферентних симпатичних і парасимпатичних нервових шляхів побудована із двох послідовно розташованих нейронів (рис. 28). Тіло першого нейрона знаходиться у ЦНС (в одному з ядер середнього, довгастого чи спинного мозку), його аксон спрямовується на периферію, але доходить лише до нервового вузла (ганглія). Тут знаходиться тіло другого нейрона, на якому аксон першого нейрона утворює синаптичні закінчення. Аксон другого нейрона іннервує відповідний орган. Унаслідок цього волокна першого нейрона називають прегангліонарними, другого - постгангліонарними, тобто еферентний шлях автономних нервів після виходу їх із ЦНС звичайно має не менше одного синаптичного контакту.
Ділянки іннервації симпатичного і парасимпатичного відділів автономної нервової системи
Симпатичні нервові волокна мають значно ширше представництво, ніж парасимпатичні. Симпатичні нерви іннервують фактично всі органи і тканини організму. Парасимпатичні нерви не іннервують скелетних м'язів, ЦНС, значну частину кровоносних судин і матку. До багатьох органів парасимпатичні волокна підходять у складі блукаючих нервів, які іннервують: бронхи, серце, стравохід, шлунок, печінку, тонку кишку, підшлункову залозу, наднирникові залози, нирки, селезінку, частину товстої кишки. Мозковий шар наднирникових залоз іннервується лише прегангліонарними волокнами. Після перерізання і переродження автономних нервів чутливість денервованих органів до відповідних трансмітерів зростає. Якщо десимпатизувати будь-який орган, що іннервується симпатичними нервовими волокнами (серце, шлунок, кишки, судини, райдужна оболонка ока тощо), то він набуває підвищеної чутливості до адреналіну і норадреналіну. Так само, якщо виконати парасимпатичну денервацію
органа, він здобуває підвищену чутливість до ацетилхоліну. Відома низка механізмів, що зумовлюють підвищення чутливості до відповідних трансмітерів: а) зростання кількості рецепторів на постсинаптичній мембрані; б) зниження активності або вмісту в тканинах ензиму, що розщеплює трансмітер. Своєрідною десимпатизацією є розвиток вікових склеротичних утворень у кровоносних судинах: у такому разі на тлі зниження впливу симпатичного нерва підвищується реакція судин на дію гормону адреналіну, викид великої кількості якого під час стресу зумовлює спазм судин. Ганглії автономної нервової системи Ганглії симпатичного відділу автономної нервової системи в залежності від їх локалізації поділяють на парні вертебральні, паравертебральні і превертебральні. Вертебральні симпатичні ганглії розташовані з обох боків хребта й утворюють два прикордонних симпатичних стовбури, отримують імпульси від нейронів, розташованих у бічних рогах спинного мозку. Аксони постгангліонарних симпатичних нейронів спрямовуються від гангліїв до периферійних органів самостійними нервовими шляхами або у складі соматичних нервів. Більшість прегангліонарних волокон симпатичної нервової системи перериваються у гангліях прикордонного стовбура. Менша їх частина проходить через прикордонний стовбур без перерви і переривається у превертебральних гангліях. Превертебральні ганглії розташовані на більшій відстані від хребта, ніж ганглії прикордонного стовбура, і дальше від органів, які ними іннервуються. Ганглії парасимпатичного відділу локалізуються всередині органів або поблизу них. Аксон першого парасимпатичного нейрона, що знаходиться у середньому, довгастому мозку чи крижовому відділі спинного мозку, доходить до органа, що ним іннервується, без перерви. Другий парасимпатичний нейрон локалізується усередині цього органа чи в безпосередній близькості від нього - у вузлі, що прилягає до нього найближче. Внутрішньоорганні волокна і ганглії утворюють сплетення, багаті на нервові клітини. Вони розташовані у м'язових стінках багатьох внутрішніх органів: серця, бронхів, середньої і нижньої третин стравоходу, шлунка, кишок, жовчного міхура, сечового міхура, а також у залозах зовнішньої і внутрішньої секреції. На відміну від симпатичного відділу, прегангліонарні парасимпатичні волокна довші, ніж постгангліонарні. Особливості волокон автономної нервової системи. Прегангліонарні волокна автономної нервової системи належать до типу В, мають тонку мієлінову оболонку. їх діаметр 2-3,5 мкм (рідше 5 мкм). Постгангліонарні волокна належать до типу С і мають діаметр не більший ніж 2 мкм. Значна їхня частина не має мієлінової оболонки. Постгангліонарні волокна характеризуються низькою збудливістю: для їхнього збудження потрібна більша сила подразнення, ніж для рухових волокон, що іннервують скелетні м'язи. Нервові імпульси поширюються ними зі швидкістю у прегангліонарних волокнах - від 3 до 18 м/с, а в постгангліонарних - від 1 до 3 м/с. Чим тонше волокно, тим менша його збудливість, триваліша рефрактерність, менші лабільність і швидкість проведення нервових імпульсів. Потенціали дії як симпатичних, так і парасимпатичних нервових волокон відрізняються більшою тривалістю, ніж потенціали дії соматичних нервових волокон. У прегангліонарних волокнах вони супроводжуються тривалим позитивним слідовим потенціалом, а в постгангліонарних волокнах - слідовим негативним потенціалом, що переходить у тривалу (до 300 мс і більше) слідову гіперполяризацію.
Медіатори автономної нервової системи
Особливості і механізми симпатичної передачі такі ж, як і в соматичній нервовій системі. В автономній нараховується більше ніж 10 трансмітерів. Ацетилхолін. Залежно від того, який трансмітер виділяється закінченнями аксонів автономних нейронів, їх поділяють на холінергічні та адренергічні. Трансмітером усіх прегангліонарних волокон є ацетилхолін, який взаємодіє з холінорецепторами постсинаптичних мембран нейронів ганглію. Розрізняють два види холінорецепторів: М і Н. М-холінорецептори втрачають чутливість до ацетилхоліну під впливом мускарину (отрута, виділена з гриба мухомора) й атропіну; Н-холінорецептори - під впливом нікотину та інших подібних гангліоблокаторів (гексоній тощо). Особливістю дії ацетилхоліну в синапсах гангліїв є те, що вона не припиняється після уведення атропіну, але зникає після дії нікотину, тобто такі синапси належать до Н-холінергічних. Холінергічними є не тільки прегангліонарні нейрони обох відділів автономної нервової системи, але й постгангліонарні парасимпатичні волокна. Постгангліонарні еферентні нейрони периферійних симпатичних гангліїв, що іннервують потові залози і забезпечують розширення судин м'язів, які працюють, також виділяють ацетилхолін. В усіх цих структурах ацетилхолін взаємодіє з М-холінорецептором. Норадреналін. Адренергічними є решта еферентних нейронів симпатичних гангліїв. У закінченнях аксонів і в синаптичних контактах, які утворені цими аксонами з непосмугованом'язовими клітинами та іншими структурами, виділяється норадреналін, що взаємодіє з адренорецепторами постсинаптичних мембран. Є два основні види адренорецепторів, з якими взаємодіють норадреналін і гормон мозкової речовини надниркових залоз адреналін: а- і р-адренорецептори. Норадреналін зумовлює значніше збудження р-адренорецепторів міокарда і менш виражену реакцію у непосмугованих м'язах бронхів. Перші називаються рр, а другі - р2-адренорецепторами. Менш істотні, але наявні відмінності і серед а-рецепторів, які також поділяють на аг і а2-адренорецептори. У більшості внутрішніх органів знаходяться два види адренорецепторів, що можуть спричинити різні чи однакові реакції. В окремих органах наявний лише один з адренорецепторів. У кровоносних судинах є і аг і а2-адренорецептори. Взаємодія симпатичного трансмітера з аг адренорецепторами в артеріальній стінці зумовлює звуження артеріол, а взаємодія з а2-адренорецепторами - їх розширення. У кишках також наявні ар і а2-адренорецептори: вплив на них гальмує скорочення непосмугованих м'язів. У міокарді і бронхах немає аг адренорецепторів, тут норадреналін та адреналін взаємодіють лише з 012-адренорецепторами. Як результат, виникає посилення серцевих скорочень і розширення бронхів. Незважаючи на те що адренорецептори взаємодіють як із норадреналіном, так і з адреналіном, їх чутливість до цих речовин різна. Так, а-адренорецептори чутливіші до норадреналіну, ніж адреналіну, а р-адренорецептори, навпаки, - до адреналіну. Це означає, що за наявності в органі обох рецепторів невелика інтенсивність збудження симпатичного нерва зумовить ефект, характерний для взаємодії норадреналіну з а-адренорецепторами. Поява в крові невеликих кількостей адреналіну може виявитися його взаємодією з р-адренорецепторами, а значної його концентрації - з а-адренорецепторами, що визначить різний кінцевий фізіологічний ефект. Норадреналін, що виділяється у синаптичну щілину, здатний впливати не тільки на рецептори постсинаптичної, але й пресинаптичної мембрани. На ній наявні обидва типи адренорецепторів. У разі взаємодії норадреналіну з пресинаптичним ос2-адренорецептором виділення трансмітера знижується, а з'єднання з р-адренорецептором посилює виділення трансмітера. Тобто спостерігається саморегулювальний механізм виділення трансмітера, безпосереднє виділення норадреналіну в синаптичну щілину шляхом екзоцитозу. Подібний механізм регулювання характерний і для відділів автономної нервової системи - симпатичного і парасимпатичного у разі подвійної іннервації органа. На пресинаптичній мембрані симпатичного синапса локалізуються М-холінорецептори і в разі взаємодії з ними ацетилхолін гальмує виділення норадреналіну. У свою чергу, мембрана нервового закінчення парасимпатичного нерва має а-адренорецептори, за участі яких норадреналін гальмує виділення ацетилхоліну. Тим самим на периферії підтримується ефект "невтручання" між зазначеними частинами автономної системи. Інші трансмітери автономної нервової системи. Крім ацетилхоліну та норадреналіну в автономній нервовій системі виявлено й інші трансмітери. У закінченнях симпатичних нервових волокон виявлено дофамін, виділення якого в синаптичну щілину відбувається під впливом нервових імпульсів. Вважають, що дофамін вступає у взаємодію з а-адренорецепторами, що локалізовані на самих пресинаптичних закінченнях, і тим самим гальмує виділення норадреналіну. У деяких відділах ЦНС, у непосмугованих м'язах кишок, матки, а можливо, і кровоносних судин, наявні рецептори до серотоніну, ефект дії якого нагадує дію трансмітера ацетилхоліну, але зберігається навіть після блокади холінорецепторів. У шлунку і кишках виявлені нейрони, збудження яких гальмує активність непосмугованих м'язів. Це здійснюється шляхом виділення закінченнями аксонів цих нейронів пуринового нуклеотиду адено-зинтрифосфорної кислоти (АТФ). Трансмітерний ефект залежить як від впливу самої АТФ, так і продуктів її розпаду - аденозину й інозину (на мембранах наявні два типи рецепторів - до АТФ і її метаболітів). Ці еферентні нейрони називаються пуринергічними. Пуринергічні нейрони є, імовірно, головними антагоністами холінергічної системи. Вони, забезпечуючи гальмування, сприяють розслабленню шлунка, кишок, стравохідного й анального сфінктерів і проштовхуванню харчового хімусу. Гістамін у низці випадків також може бути трансмітером, тому що в деяких тканинах виявлені специфічні Нг та Н2-гістамінорецептори. Гістамін є біологічно активною речовиною широкого спектру дії. Установлено, що значно поширений у синапсах ЦНС гальмівний трансмітер гама-аміномасляна кислота (ГАМК) гальмує проведення збудження у зірчастому ганглії, але полегшує передачу його у верхньому шийному, нижньому брижовому сплетеннях і в гангліях сонячного сплетення
8.4. Рефлекси автономної нервової системи
Нейрони автономної нервової системи беруть участь у здійсненні багатьох рефлекторних реакцій, які можуть виникати за умови подразнення як інтерорецепторів, так і екстерорецепторів. Ознакою того, що ці рефлекси належать до автономних, є надходження імпульсів до еферентного периферійного органа з ЦНС по симпатичних чи парасимпатичних нервах.
Рефлекси гангліїв автономної нервової системи
Функціональна характеристика гангліїв Автономні ганглії відіграють важливу роль у розподілі і поширенні нервових імпульсів, що проходять через них. В основі цього - дві структурні особливості гангліїв: 1) кількість нервових клітин у кілька разів (у верхньому шийному вузлі - у 10 разів, у війковому вузлі - у 2 рази) більша, ніж кількість прегангліонарних волокон, що надходять до ганглію; 2) кожне з пресинаптичних волокон сильно розгалужується, утворюючи синапси на багатьох клітинах ганглію. Тому нервові імпульси, що надходять по прегангліонарному волокну до ганглію, можуть впливати на велику кількість гангліонарних нейронів і, отже, на ще більшу кількість м'язових і залозистих клітин того органа, який іннервується (явище дивергенції). Таким чином досягається розширення зони впливу прегангліонарних волокон. На кожному гангліонарному нейроні є синапси, утворені великою кількістю прегангліонарних волокон. Це створює умови для конвергенції -просторової сумації нервових імпульсів. Надходження одного нервового імпульсу до гангліонарного нейрона ніколи не викликає його збудження -для цього обов'язково необхідна сумація (як просторова, так і часова). Тому в природних умовах дивергенція і конвергенція відіграють істотну роль у забезпеченні відповідного регулювального впливу на функції внутрішніх органів. Однобічне проведення нервових імпульсів у міжнейронних синапсах, перекриття зон впливу окремих прегангліонарних волокон, які входять у ганглій, наявність часової і просторової сумації й оклюзії показує, що нейрони і синапси гангліїв автономної нервової системи мають такі ж властивості, як і нейрони та синапси ЦНС. В інтрамуральних гангліях парасимпатичної нервової системи явище дивергенції виражено менше, ніж у симпатичній, тут часто зустрічаються прямі контакти прегангліонарного нейрона з одним постгангліонарним нейроном. Рефлекси, що замикаються на рівні гангліїв автономної нервової системи, називають рефлексами її метасимпатичного відділу. Ганглії автономної системи є винесеними на периферію рефлекторними центрами з елементами інтегративної діяльності. У них є усі нейрони, необхідні для виконання рефлекторного переключення (аферентні, еферентні, вставні, у тому числі й гальмівні). Не виняток й інтрамуральні ганглії та нервові сплетення, що локалізовані в порожнистих органах. Ці ганглії є частиною еферентного шляху парасимпатичної нервової системи. Але одночасно до них підходять нервові клітини від внутрішньоорганних рецепторів. Усі ці нервові клітини утворюють між собою синаптичні контакти, так що вже у самому ганглії можлива передача впливів від чутливого нейрона до еферентного. Будова інтрамуральних гангліїв нагадує типові нервові центри. Кожен нейрон оточений великою кількістю клітин нейроглії. Тут є структури, що вибірково пропускають до нейрона з крові лише визначені речовини, цим вони нагадують гематоенцефалічний бар'єр. Таким чином, нейрони ганглія, подібно до нейронів мозку, захищені від безпосереднього впливу речовин, що циркулюють у крові. Серед інтрамуральних еферентних нейронів є не тільки холінергічні, але й адренергічні, а також пуринергічні, серотонінергічні, дофамінергічні і, очевидно, гістамінергічні, пептидергічні і ГАМК-ергічні. Усе це створює можливість широкого діапазону регулювальних впливів. "Місцеві" периферійні рефлекси, що реалізуються інтрамуральними автономними гангліями, регулюють роботу серця, перистальтику кишок, здійснюють взаємозв'язок різних відділів шлунка і деяких інших органів. Нейрони, що входять у ці ганглії, їх відростки, синаптичні закінчення формують внутрішньоорганні рефлекторні структури, які регулюють роботу органа шляхом внутрішньоорганних периферійних рефлексів. Серед структур метасимпатичної нервової системи є пейсмекерні клітини, які володіють здатністю до мимовільної деполяризації, що забезпечує ритм скорочення непосмугованих клітин органа. Ця активність коригується імпульсами власної аферентації у залежності від функціонального стану органа та стану його окремих частин. Імпульси, що надходять до органа прегангліонарними волокнами парасимпатичних нервів, вступають у взаємодію з імпульсами, що забезпечують процеси внутрішньоорганного рефлекторного регулювання. Характер відповідної реакції органа визначається результатом зазначеної взаємодії. Тому ефект подразнення прегангліонарних волокон не буває однозначним. На органи, у яких виявлено інтрамуральні рефлекторні механізми регулювання, прегангліонарні парасимпатичні волокна можуть здійснювати (залежно від функціонального стану органа) як збуджувальний, так і гальмівний вплив. Вони можуть запускати чи припинювати, підсилювати або послаблювати ту чи іншу функцію органа, здійснюючи різноманітні регуляторні впливи, необхідні для підтримання нормальної поточної діяльності і забезпечення гомеокінезу. На характер "місцевих" рефлексів може впливати і симпатична нервова система. Але, на відміну від парасимпатичного, симпатичний відділ автономної нервової системи за різної сили подразнення здійснює на органи однотипні впливи. Волокна симпатичної системи, що підходять до органа, - постгангліонарні. Вони закінчуються безпосередньо на клітинах органів і тканин. І хоча імпульси, що надходять по них, майже не вступають у взаємодію з елементами внутрішньоорганної рефлекторної дуги, проте трансмітери симпатичних нервів, які виділяються поблизу структур метасимпатичного відділу, можуть впливати на виділення трансмітера в них. Таким чином, еферентні інтрамуральні нейрони являють собою загальний кінцевий шлях для імпульсів внутрішньоорганного і позаорганного походження. Наявність "місцевих" механізмів нервового регулювання функцій внутрішніх органів, що здійснюється за допомогою периферійних рефлексів гангліями автономної нервової системи, внутрішньоорганними і позаорганними, має велике фізіологічне значення. Насамперед це звільняє ЦНС від необхідності переробки надлишкової інформації, що надходить від внутрішніх органів. Крім того, периферійні рефлекси збільшують надійність регулювання фізіологічних функцій цих органів. Таке регулювання є базовим і спрямоване на збереження гомеостазу. Водночас воно в разі потреби може легко коригуватися вищими рівнями автономної нервової системи і гуморальними механізмами. Це регулювання може здійснюватися навіть і після переривання зв'язку органів із ЦНС. Рефлекси симпатичних гангліїв. Периферійні рефлекси здійснюються і превертебральними гангліями, однак, імовірно, не всіма. Так, до цього часу не виявлено такої діяльності у шийних симпатичних гангліях. Еферентні нейрони автономних гангліїв отримують імпульси як з ЦНС (через прегангліонарні автономні волокна), так і від внутрішньоорганних рецепторів, утворених дендритами аферентних нейронів. їх рефлекси мають багато спільного, проте відрізняються від рефлексів метасимпатичного рівня. їх істотною відмінністю є значне поширення рефлекторної відповіді, коли завдяки наявності явищ конвергенції і дивергенції у відповідь втягуються віддалені ділянки органів чи навіть органи іншої функціональної системи (наприклад, кровообігу і травлення). Спінальні рефлекси
На рівні спинного мозку замикаються рефлекторні дуги багатьох автономних рефлексів. Характер рефлекторної відповіді визначається наявністю нервових центрів симпатичного (груднинно-поперековий) і парасимпатичного (крижовий) відділів автономної нервової системи. Спинномозковий відділ має риси сегментарної (метамерної) організації. Це проявляється у тому, що найбільш чітке переключення чутливих входів на еферентні відбувається у межах конкретного сегмента спинного мозку. Хоча наявні і зони перекриття сусідніх сегментів, але відповідь на подразнення сусідніх корінців менш виражена. Найпоказовіші в цьому плані рефлекси серцево-судинної і травної систем, видільних органів (кардіо-кардіальні, кишково-кишкові, видільні). Характер рефлекторної відповіді, яка замикається на рівні спинного мозку, залежить ще й від вставних нейронів. Інтернейронний апарат спинного мозку забезпечує взаємодію рефлекторних шляхів як всередині автономної системи, так і між нею і соматичною нервовою системою. Завдяки цьому відбувається широке залучення різних внутрішніх органів у рефлекторну відповідь. Спінальні центри регулювання автономних функцій. На рівні останнього шийного і двох верхніх грудних сегментів спинного мозку знаходяться нейрони, що іннервують три непосмуговані м'язи ока: м'яз-роз-ширювач зіниці, очноямкову частину кругового м'яза ока та один із м'язів верхньої повіки. Ділянка спинного мозку, від якого йдуть нерви до цих м'язів, називається спіноциліарним центром. Нервові волокна від цієї ділянки проходять у складі симпатичного нерва до верхнього шийного симпатичного вузла, де починається другий нейрон, який закінчується в очних м'язах. Подразнення цих волокон викликає розширення зіниці (мідріаз), розкриття повікової щілини і випинання очного яблука (екзофтальм). Ураження названих сегментів спинного мозку чи перерізання симпатичних нервів призводять до розвитку синдрому Горнера: звуження зіниці (міоз), звуження повікової щілини і западання очного яблука (ендофтальм). У верхніх грудних сегментах спинного мозку розташовані нейрони, що входять до складу центра, який регулює роботу серця і стан кровоносних судин. Тут також наявні нейрони, які іннервують бронхи. В усіх грудних і верхніх поперекових сегментах спинного мозку розташовані нейрони, що іннервують потові залози та органи травлення. Ураження окремих сегментів зумовлює припинення потовиділення у ділянках тіла, що втратили симпатичну іннервацію. У крижовому відділі спинного мозку розташовані спінальні центри рефлексів сечовипускання, дефекації, ерекції та еякуляції. Руйнування зазначених центрів спричиняє статеве безсилля, нетримання сечі і калу. Порушення сечовипускання і дефекації відбувається унаслідок паралічу сфінктерів сечового міхура і прямої кишки. Автономні рефлекси дуже численні. Для медичної практики мають велике значення вісцеро-вісцеральні, вісцеро-дермальні, дермато-вісцеральні, вісцеро-соматичні і сомато-вісцеральні рефлекси. Вісцеро-вісцеральні рефлекси викликаються подразненням рецепторів, що розташовані у внутрішніх органах, і припиняються у разі зміни діяльності останніх. Ці рефлекси можуть виникати і закінчуватися в органах однієї функціональної системи (наприклад серцево-судинної) або бути міжсистемними. До вісцеро-вісцеральних рефлексів належать рефлекторні зміни серцевої діяльності, тонусу судин, кровонаповнення селезінки внаслідок підвищення або зниження тиску в аорті, каротидному синусі чи легеневих судинах, рефлекторна зупинка серця у разі подразнення органів черевної порожнини тощо. Вісцеро-дермальні рефлекси виникають у разі подразнення внутрішніх органів і виявляються змінами потовиділення, електричного опору (електропровідності) шкіри і шкірної чутливості на обмежених ділянках поверхні тіла, топографія яких різна залежно від того, який орган подразнюється. Дермато-вісцеральні рефлекси полягають у тому, що під час подразнення певних ділянок шкіри виникають судинні реакції і зміни в діяльності конкретних внутрішніх органів. У разі вісцеро-соматичних рефлексів подразнення рецепторів внутрішніх органів спричинює зміну активності скелетних м'язів і, як наслідок, активування або гальмування м'язових скорочень. Сомато-вісцеральні рефлекси виявляються зміною діяльності внутрішніх органів унаслідок скорочення скелетних м'язів, наприклад, під час фізичних навантажень. Названі автономні рефлекси використовуються у практичній медицині. Наприклад, для оцінки функціонального стану судин досліджують рефлекторні зміни їх тонусу під час механічного подразнення шкіри. У багатьох практично здорових людей у разі механічного подразнення шкіри тупим предметом (неврологічним молотком) виникає місцеве звуження артеріол, що проявляється у вигляді нетривалого збліднення ділянки шкіри, яка подразнюється (білий дермографізм). За більш високої чутливості з'являється червона смуга розширених шкірних судин, облямована блідими смугами звужених судин (червоний дермографізм), а за наявності дуже високої чутливості - смуга ущільнення шкіри і її набряк.
|