Потребность и влечение
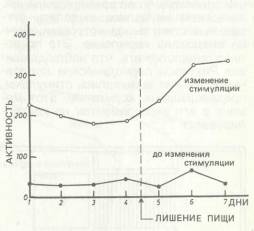
Промежуточную позицию в спорах об инстинктах занимал Вудвортс [R. Woodworth, 1918]. С одной стороны, полемизируя с Мак-Дауголлом, он выступал против абсолютизации инстинктов как последнего мотивационного основания детерминации поведения. С другой, не соглашаясь с бихевиористским ассоцианизмом, он оспаривал объяснительную ценность чистых S — R-связей. Он вставил между стимулом и реакцией еще один детерминант — состояние организма — и обогатил уравнение поведения S — R членом О (организм): S — О — R. Раз существует организмическое состояние нужды, следует, как предложил еще Шеррингтон, различать подготовительные и консумматорные реакции (конечные действия). Подготовительные реакции могут в весьма значительной степени определяться внешними стимулами, на консумма-торных, напротив, сказывается влияние внутренних стимулов, влечений (drive), которые приводят поведение к завершению, к удовлетворению, к покою. Рассматривая поведение «динамически», Вудвортс указывал на возможность приобретения «механизмами» поведения (структурный компонент) характера стабильного влечения (т. е. превращение в мотивационный компонент). В результате под стимулами начали понимать не только внешние воздействия на организм, но и внутренние возникающие в самом организме и оказывающие на него свое действие стимулы. Фрейд еще в 1895 г. отмечал специфику внутренних раздражителей, как таких, от которых организм не может спастись бегством, в физиологии начался поиск способов регистрации внутренних стимулирующих различные формы поведения раздражителей. Для голода и жажды Кенноном с сотрудниками была разработана локальная теория мотивации [W. В. Cannon, A. L. Washburn, 1912]. При помощи проглатываемого резинового шарика, который в желудке надувался, измерялись желудочные сокращения. Они коррелировали с ощущениями голода. Внутренними организмическими стимулами, ответственными за чувство жажды, считались ощущения пересыхания слизистой оболочки рта. Под напором развернувшихся в последующие десятилетия интенсивных исследований [см.: R. Bolles, 1967; 1975] локальная теория мотивации рухнула. Например, оказалось, что собаки, которых поили «не по-настоящему» (выпитая вода выводилась наружу через вставленную в пищевод трубку раньше, чем достигала желудка), выпивали чрезвычайно большое количество воды, хотя их полость рта постоянно была влажной. Регуляция приема пищи и жидкости оказалась необычайно сложной, и до сих пор ее физиологические механизмы полностью не раскрыты. Наряду с периферическими органами (пищевой тракт, желудок, кишечник, печень, соматические клетки, артериальное и венозное кровообращение) в ней принимают участие и мозговые центры как центральные интегративные механизмы [S. Balagura, 1973]. С физиологической точки зрения более сбалансированную теорию влечений предложил Морган [С. Т. Morgan, 1943], назвав ее теорией «центральных мотивационных состояний»-При различных видах мотивации, как, например, голод, жажда или половое влечение, за счет внешних и внутренних раздражителей, гормональных факторов и факторов опыта возникает специфическое для каждого мотива «центральное мотивационное состояние», которое характеризуется следующими признаками: 1. Стойкость. Длительность состояния превышает время появления инициирующих условий и последующего поведения. 2. Общая активность. Ее уровень постепенно возрастает. 3. Специфическая активность. Вызывает специфические формы поведения, не зависящего от конкретных условий ситуации. 4. Подготовленность. Повышение готовности организма к консумматорному поведению, определяемому соответствующими средовыми условиями. Все эти признаки, считает Морган, нельзя объяснить воздействием внешних или внутренних стимулов. И хотя центральные мотивационные состояния детально изучены физиологически, но на сегодняшний день мало изучена их связь с последующим протеканием поведения, что делает это понятие малопригодным для психологического объяснения поведения. В другом направлении исследований, связанном в первую очередь с именем Рихтера, за индикатор периодических колебаний влечений была принята общая активность подопытных животных, которая, очевидно, зависит от циклических колебаний потребностей, необходимых для поддержания организма в состоянии равновесия (гомеостаза). День за днем в барабанах для бега автоматически регистрировалась активность животного. Интерпретация зарегистрированных колебаний активности строилась на трехчленной схеме детерминации: потребность через внутреннюю стимуляцию вызывает влечение, or которого линейно зависит повышение активности. Долгое время считалось, что физиологические индикаторы потребностных состояний свидетельствуют о наличии соответствующего влечения и что последнее, пока оно не удовлетворено, ведет к повышению общей активности. Вначале даже считали, что при помощи принципа гомеостаза можно объяснить любое поведение [R. В. Raup, 1925]. Однако вскоре оказалось, что выводы о наличии влечения, основаны ли они на предварительных индикаторах потребности (need) или на последующем повышении активности, слишком поспешны и прямолинейны. И в этом случае все оказывается значительно сложнее. Повышение и понижение активности изголодавшихся крыс гораздо сильнее зависят от условий внешней стимуляции, чем это кажется на первый взгляд. Это можно пояснить на примере эксперимента Кэмпбелла и Шеффилда [В. Campbell, F. Sheffield, 1953]. Они держали крыс семь дней в специальных клетках, опора которых позволяла регистрировать каждое движение животного. Экспериментальное помещение было темным и звукоизолированным, вентилятор создавал равномерный звуковой фон, маскирующий остаточные шумы. Первые четыре дня пищи было достаточно, последние три дня ее не давалось вовсе. Раз в день экспериментатор входил на 10 мин в помещение, включал свет и выключал вентилятор. Активность регистрировалась как во время изменения стимуляции, так и за 10 мин до этого. На рис. 4.1 приведен график изменения активности на протяжении семи дней. В отсутствие изменений стимуляции активность сохраняется на одном и том же низком уровне, хотя начиная с четвертого дня голод возрастает. В периоды изменения стимуляции, напротив, активность устойчиво выше и увеличивается с возрастанием голода.
Рис. 4.1. Средние изменения активности, измеренной в 10-минутные интервалы до или во время ежедневного изменения стимуляции в сытом состоянии (1 — 4 дни) и при возрастающем голоде (5 — 7 дни) [В. Campbell, F. Sheffield, 1953, p. 321]
Эти данные противоречат предположению Рихтера об автоматическом возрастании активности с обострением потребностного состояния. Что явно возрастает, так это готовность реагировать на внешние стимулы. В какой-то мере приведенные данные подтверждают рассуждения Моргана об усиливающемся общем мотиваци-онном состоянии. В последующих экспериментах Шеффилд и Кэмпбелл [F. Sheffield, В. Campbell, 1954] показали, что рост активности в период голодания особенно выражен, если в предыдущие дни изменение стимуляции происходило во время кормления. Животные научались выделять сигнальные стимулы, за которыми обычно следовало кормление. Это позволяет предположить, что наблюдавшиеся Рихтером периодические колебания влечения вызывались стимулами, предвещавшими кормление, этот момент в его экспериментах не контролировался.
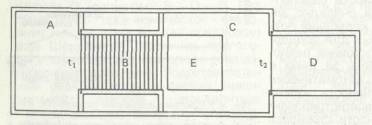
Рис. 4.2. Колумбийский проблемный ящик с препятствием для измерения уровня активности, вызываемой конкретным влечением [Т. Jenkins, L. Warner, С. Warden, 1926, p. 366]
Тем не менее к перспективе выявить и измерить влечение через наблюдаемые показатели исследователи относились оптимистически. Регистрация общей активности была сопряжена с трудностями интерпретации, поскольку каждый раз оставалось под вопросом, с каким конкретно влечением мы имеем дело. Ведь общая активность не целенаправлена соответственно конкретному влечению. Поэтому шагом вперед явилось измерение конкретной и целенаправленной активности при помощи нового экспериментального аппарата: колумбийского ящика с препятствием. Его чертеж представлен на рис. 4.2' Чтобы добраться до объекта, побуждающего влечение, животное, помещенное во входную камеру (А), должно пробежать по решетке, через которую пропускается электрический ток (В). Дверца, ведущая к ней (t,), открывается экспериментатором. Когда животное преодолевает препятствие, оно попадает в переднюю часть камеры с подкреплением (С) и наступает на пластинку (Е), открывающую дверцу (t2) в заднюю часть этой камеры (Д), где собственно и находится специфический объект конкретного влечения (пища, вода или партнер по полу). Вначале животным давалась предварительная тренировка, чтобы они ознакомились с устройством при наличии объекта влечения, и лишь во время последних тренировочных пробежек через решетку пропускался ток. В основных экспериментах варьировалась продолжительность лишения возможности удовлетворить специфическую потребность (депривация) и затем в течение 20 мин регистрировалось, сколько раз животное стремится преодолеть боязнь решетки с электроразрядами, чтобы добраться до привлекающего объекта. Таким путем рассчитывали не только определить зависимость интенсивности или настоятельности отдельных влечений от продолжительности деп-ривации, но и сравнить различные виды влечений. Как видно из рис. 4.3, жажда, по-видимому, быстрее достигает своей интенсивности, чем голод, а голод — быстрее, чем половое влечение у самцов. Конечно, число попыток преодолеть препятствие по ряду причин не может служить надежным индикатором интенсивности влечения. Например, во время предварительной тренировки неконтролируемые факторы могли привести к различным результатам научения. На результатах может также сказаться изменение длительности наблюдения. Вряд ли можно решить, какая продолжительность наблюдения наиболее валидна для измерения интенсивности данного влечения. Прежде всего в опыте систематически не контролировалась привлекательность объекта. Между тем, как теперь хорошо известно, этот мотивирующий фактор может активировать поведение независимо от потребностного состояния. Кроме того, при каждом контакте с объектом влечения, каким бы мимолетным этот контакт ни делал экспериментатор, возникают различные проявления консумматорной активности, которые нельзя считать полностью сопоставимыми между собой.
Рис. 4.3. Зависимость от длительности депривации частоты, с которой крысы преодолевали в 'ящике решетку,- пропускавшую ток, с тем, чтобы добраться до объекта специфической потребности [Т. Jenkins, L. Warner, С. Warden, 1926]
|